History of PGR conservation
Motivation
The current efforts to collect, preserve, and utilize plant genetic resources are the result of a long historical process. It is necessary to know about the history of plant genetic resources to understand the current discussion at the interface of politics and science of plant genetic resources.
Although the interest in the conservation plant genetic resources began at about the same time as modern scientific plant breeding, i.e. the beginning of the 20th century, the key organisations and international frameworks that shape the current system of PGR conservation were established after the second world war and also after the breakdown of the wall in 1989.
The modern phase of the plant genetic resource conservation movement began when it was realized that the worldwide introduction of modern high-yielding, high-input varieties of wheat and rice, which is called the Green Revolution1, led to the rapid disappearance of local land races of these crops and of crop wild relatives.
1 The period of agricultural development ranging from the 1940s to 1970s
At about the same time, a system for the collection and preservation of genetic resources was established, which evolved into the system of the Consultative Group of International Agricultural Research (CGIAR)2 institutes. In this system, each institute has a mandate for a particular crop or agroecosystem and promotes research and the development of new varieties using state of the art breeding technologies.
Another important development was the Convention on Biological Diversity (CBD)3 in 1992, which led to a formal definition of the term 'biodiversity'. The CBD also is the foundation for international regulations of exchange of the genetic material between countries and it assigned the task of preserving and taking stewardship of genetic resources to each country. In the context of plant genetic resources, the International Treaty on Plant Genetic Resources and the Nagoya Protocol are an outcome of the CBD that every breeder has to know and understand, because these international regulations have a huge influence on the utilization of plant genetic resources in breeding.
3 www.cbd.int
In the following, we will take a historical approach to show that the current situation results from historical developments. The narrative of events may come across as somewhat long and dry (and even boring), but putting them into a context is important to understand that a quick action was necessary to prevent the loss the results of 10,000 years of crop genetic diversity within a few decades.
Learning goals
Plants that changed the world
Since the era of plant domestication, some crop species expanded their distribution range and became important crops, both globally and locally, while other crops remained locally restricted for ecological or historical reasons.
Examples of globally successful crops are wheat and barley, which were domesticated in the Near East and were cultivated in large regions of Europe by the end of the neolithic age. A similar expansion was experienced by maize, which was domesticated in Mexico, but could be found at the time of the conquest throughout South, Central and North America. The third major crop of today, rice, has experienced a similar range expansion throughout Asia. It should be noted that the expansion of these crops was fairly slow and took hundreds to thousands of years. Furthermore, it was restricted to geographic regions (e.g, barley and wheat to the old world and Asia, rice to East and South Asia, maize to the Americas). The combination of archeological findings and genetic studies allows to model the geographic spread of crops and it shows that for example the spread of barley in Europe was associated with the expansion of agriculture (Figure 1).
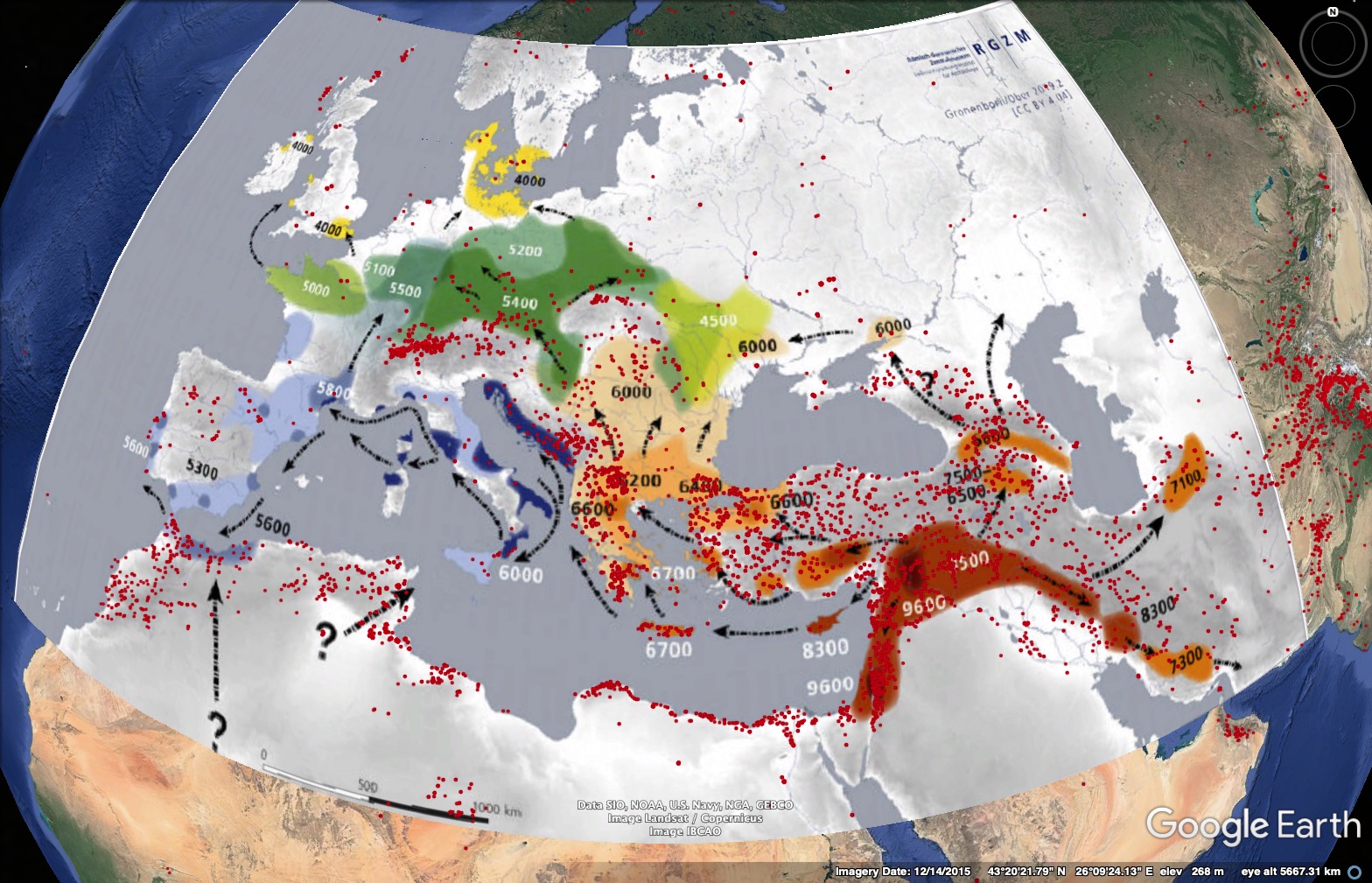
However, since the discovery of the Americas by Columbus, within 500 years, the exchange of crop plants was much faster and one consequence was that the rapid global expansion of a small number of crop plant species literally transformed the world. This transformation that was not only influenced by plants, but also by animals and associated diseases, has been called the Columbian exchange (Crosby, 1972). It has wide ranging influence until today and shows the importance of plant genetic resources and their power in transforming local, traditional agriculture and food security of human societies.
Beginning with the modern era and the age of great discoverers, many other plants were distributed around the world in a short time, where they had a huge impact on local agriculture and society. These plants were mostly exported to regions with a similar climate to their region of origin, and there was little improvement and breeding to adapt them to their new environments. Some plants were extremely important in alleviating famine and disease, others provided huge profits to their growers: Sugar cane, quinine, tea, cotton, potato, timber, wine, rubber and tobacco (Hobhouse, 2006). These plants were central to the development of colonialism and the post-Colombian imperialism that affected the lives of hundreds of millions of people and continues to leave its footprint until today. Not only the colonial cash crops were important, but also field crops like maize and potato, which affected the life of small scale farmers outside their original distribution area.
Introduction of maize into Europe
As an example of the transformative power of the a Colombian exchange in the context of plant genetic resource, the introduction of maize into Europe is presented.
The analysis of historical patterns of crop distribution is relevant for the question of where valuable genetic diversity of a crop species can be expected and utilized for plant breeding. The historical pattern of migration has been studied for several crop species, such as maize and potato. For maize, the 'migration' of different subtypes of maize during the last 500 years is shown in Figure 2 and based on historical and genetic analyses.
Over the last years, it has been hypothesized by historians that maize was introduced only once by Columbus into Europe. This notion was challenged by geneticists who know the difference between the 'flint' and 'dent' types of maize, and also the different degrees of adaptation of European maize varieties to the warmer climates of the South and the colder climates of Northern Europe. Several introductions were postulated (Figure 2). This hypothesis was investigated using genetic markers. A representative set of American (North, Central, South) and European maize lines was selected for genotyping with RFLP (restriction fragment length) and SSR (simple sequence repeats) molecular markers. A phylogenetic tree calculated with these markers indicates that European maize lines are strongly differentiated by latitude, i.e., into Northern and Southern European groups. (Figure 3 A).
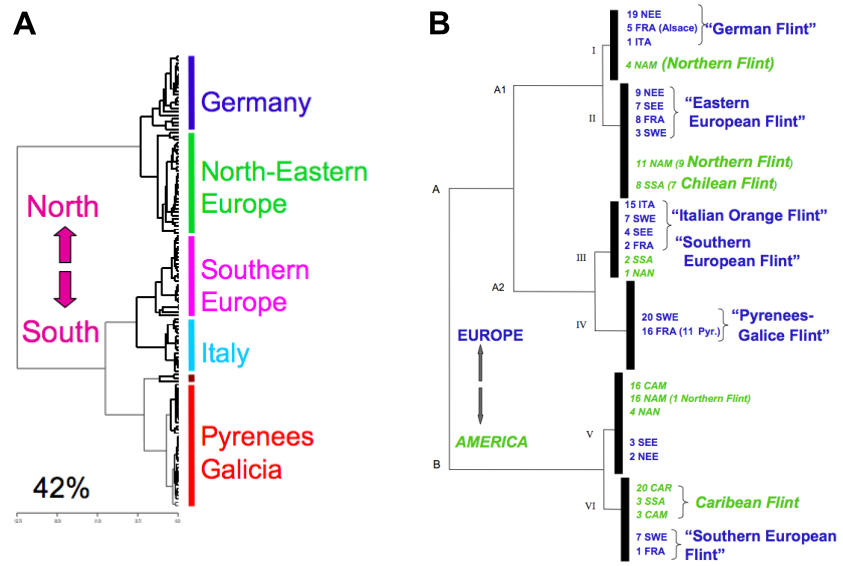
To understand the origin of European maize landraces, a core collection of 100 populations was generated that contained 93% of the variation of a large collection of 2,000 populations based on 23 RFLP loci, phenotypic information and passport data (Gauthier et al., 2002). A somewhat larger set of 139 European accessions was analysed together with 89 American maize populations using 29 RFLP markers to identify genetically similar accessions from Europe and America (Rebourg et al., 2003)
Figure 3 B shows that there are similarities between
- Southern Spain and Caribbean populations, consistent with an initial introduction by Columbus
- Northern European populations and American Northern Flints, also with populations from South Chile
- Italian and South American populations
- Furthermore, there was an 'original' group in the Pyrenees without similarity to American populations.
In summary, the molecular results suggest a strong contribution of Northern Flint populations to the adaptation of maize to Northern Europe:
- as such in North Eastern regions
- through hybridization with late populations, to generate traditional varieties at intermediate latitudes (Pyrenees, Galicia, etc).
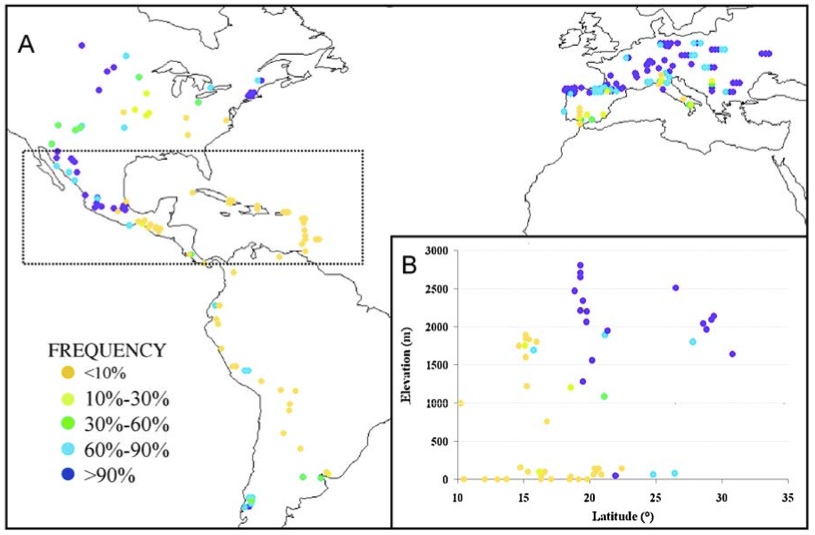
Given the environmental differences between the Americas and Europe, i.e., with respect to daylength and annual season, the introduced maize populations had to adapt to European cultivation conditions and thereby leave a strong footprint of adaptation in genes controlling adaptive traits. For example, the geographic distribution of a genetic marker (a transposable element of the MITE family), which is linked to the trait early flowering, which is controlled by the Vgt1 locus, indicates that after the introduction of the maize landraces into Europe, local selection caused an adaptation to European conditions as indicated by an increase in allele frequency of the allele for early flowering (Figure 4).
This genetic interpretation is consistent with an historical analysis. For example, the first picture of maize in a European book shows the typical tillering pattern present in Northern Flint (Figure 5).

The German herbalist Jerome Bock described in 1539 the typical pattern of maize cobs that refers to the ear morphology of the Northern Flints. In conclusion, Northern Flint populations was broadly spread in Germany (1539) and cultivated in gardens almost everywhere.
The genetic studies lead to the conclusion that the introgression of maize into Europe was not a slow migration process that started with introgressions into Southern Spain and a subsequent spread into Northern Europe, which was frequently assumed in the historical sciences. Instead, not only tropical varieties were introduced by Columbus, but also Northern Flint populations from North America were introduced early and played a key role for adaptation to Northern temperate climates, either as a 'pure' flint lines or by outcrossing with tropical material.
In conclusion, the research shows that a high level of genetic diversity can be expected in European maize landraces as well as (genomic) signatures of local adaptation. Similar patterns of introduction and expansion have been described for other crops such as rice.
Historical development and collection expeditions
After the Columbian Exchange, the commercial value of plants was recognized by the growing commercial enterprises and imperial states that were seeking to extract as much profit from the colonies as possible. With the recognition of the value of genetic resources, various expeditions aimed at solely collecting genetic resources have been initiated since the beginning of the 20th century. They differed by their purpose to broadly collect as many potentially useful species as possible, or to increase the sampling breadth of certain crops. During the imperial age that followed the discovery of the Americas, the collection and exchange of plants was increasingly seen as a means to strengthen one's own empire. All large empires conducted such collection trips that led to the growing importance of some plants in the imperial trade.
In the following, a few examples of well documented expeditions are presented.
Paul Preuss in Latin America (1899/1900)
The botanist Paul Preuss was the head of a Botanical Garden in Victoria, Cameroon (then a German colony) and undertook a year-long expedition to Surinam, Guyana, Trinidad, Grenada, Venezuela, Ecuador, Nicaragua, Salvador, Guatemala, Mexico, Cuba and Jamaica. The journey was financed by a lottery and by various industrial and plantation companies. One goal was to strengthen agriculture of the German colony in Cameroon by introducing new crop plants. Therefore, better varieties of cocoa, cinnamon and vanilla were searched for later introduction into the German colonies.
Another goal was to break up existing monopolies because different empires controlled the trade with certain plants or their products. For example, Peru balm was produced exclusively in El Salvador. The balm is a secretion of the tree Myroxylon balsamum and was used for medicinal and cosmetic purposes. By introducing the plant to Cameroon, the goal was to break the exclusive production of the balm in El Salvador.
Preuss collected about 20,000 seeds and afterwards distributed them to various institutions and companies in the German colonies in Africa. In addition to the exotic tropical crops, food crops such as maize and various beans were also collected.
Origin of scientific plant breeding
The history of plant genetic resources needs to be considered in the context of modern scientific plant breeding because the need for the introduction of new, exotic and genetically diverse material into breeding populations was recognized early, which led to collection expeditions aimed at preserving or obtaining novel genetic diversity.
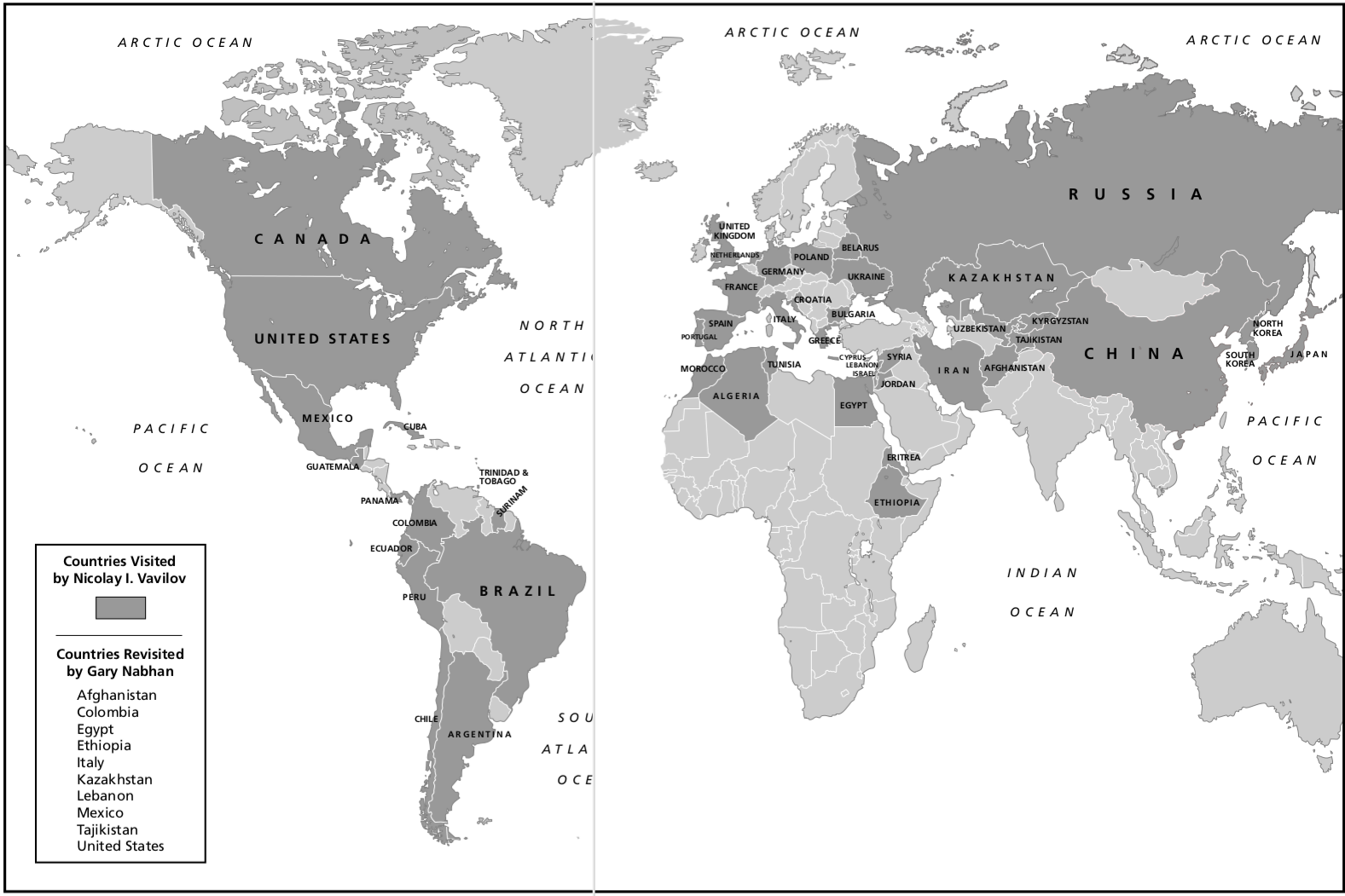
Therefore, a short history of scientific plant breeding is provided to put the history of plant genetic resources into context. In the mid 1860's Gregor Mendel published his research on the plant hybridization (crosses), which was re-discovered in 1900. This rediscovery was rapidly integrated into formal breeding in Germany, UK, USA, Sweden and Russia. One early challenge was to map genetic diversity and use it in breeding. In parallel, the Russian geneticist Nikolai Vavilov went on collection expeditions between 1900 and 1920 and visited many countries worldwide to collect 'primitive' forms (landraces) and wild relatives of crops. These collection expedition and the subsequent scientific analysis led to his theory of centers of domestication, which in his honor were also called Vavilov centers.
Between 1920 and 1930, plant breeding developed into a formal science because of the theoretical developments in population and quantitative genetics. In particular the foundational paper of R. A. Fisher established the field of quantitative genetics because he developed a theory that reconciled Mendelian genetics (inheritance of individual genes with major phenotypic effects) with Darwin's evolutionary paradigm of gradual phenotypic changes over time.
During this time, early versions of plant variety protection (PVP) developed, which regulated the marketing of seeds and fostered the development of seed companies. Until the late 1930's, the USA, Germany, Sweden and the Sovjet Union became nations in plant breeding. In the Sovjet Union, Nikolai Vavilov was the leading plant geneticist. However, he had a powerful opponent, Trofim Lysenko, who was a protege of Josef Stalin. Lysenko proposed that plant traits are acquired characteristics and not inherited, which fitted to the political views of Stalin that workers can escape capitalist oppression by recognizing their situation and starting a revolution. In addition, science Lysenko was born into a peasant family and Vavilov originated a wealthy family of merchants in St. Petersburg, Lysenko also was closer to the communist ideal of a peasant/worker vs. burgeois science, as which genetics was considered 4.
4 A very good documentary about this conflict can be found on YouTube at https://www.youtube.com/watch?v=K6FOFuo0bqs . Unfortunately, it is in German only, but English subtitles can be generated.
During this time, scientific plant breeding was mainly conducted at Kaiser-Wilhelm-Institutes (KWI). In 1939, the National Socialist leaders (with support of some scientists) realized that genetic material is required for both scientific research and also food security. For this reason, German KWI scientists followed the German troops behind the lines during the attack of the German army in neighboring countries and collected genetic material, which was sent back to Germany. A key event in the second world war was the defeat of the German army in Stalingrad, where a large number of German soldiers were lost, and which led to the advance of the Red Army. This lead to a more systematic collection of genetic resources, which is outlined below.
The development of plant breeding in Germany
The following presentation shows the relationship between both domains by the history of plant breeding in Germany.
At the end of the 19th century, the era of scientific plant breeding began, mainly by using the approach of mass selection, and in some crops by comparing the performance of single elite lines extracted from populations. Although some breeders were extremely successful such as Ferdinand von Lochow who developed new rye varieties, the main improvements in agricultural productivity at that time were achieved by changing various aspects of plant production (fertilization, ploughing, etc.) rather than plant breeding.
Im some environments modern varieties did not produce a higher yield than old adapted land races. For this reason, the interest in genetic diversity increased. At an Agricultural Conference in Vienna in 1890, von Proskowetz and Franz Schindler stated the importance of land races in plant breeding: 'What is the relationship in value between land races of agricultural crops and the so-called breeding lines?'. They postulated to examine the breeding value of land races.
With the re-discovery of the Mendelian rules in 1900, genetics became an important theoretical concept in plant breeding. For example Hermann Nilsson-Ehle in Sweden was developed in 1907 the so-called Panzerweizen by crossing a high-yielding English Squarehead with a strongly cold- and frost-tolerant Swedish land wheat. The result was a combination of both favorable traits.
Mass selection remained the predominant method of breeding until after the first World war, both in companies, most of which were located in North Germany, as well as in public breeding institutes that were founded early in the 20th century. The state breeding institute in Hohenheim was founded in 1905, in Weihenstephan near Freising in 1902 and the Badian institute Hochberg in 1908.

The plant geneticist Erwin Baur spoke in 1913 at the meeting of the German Society of Agriculture (DLG) about the loss of old varieties by the success of modern plant breeding and he demanded the protection of old races in order to maintain future breeding progress. Baur himself and some others as well as private breeders established at that time large collections of German land races. There were no considerations of the broader geographical context of crops. On the other hand, the potato, whose origin was known to be located in South America, played a very important role in the idea that resistance genes can be crossed into modern varieties from wild relatives. This led to a growing interest in the collection of resources from the area of domestication.
At the end of the 19th century, the commercialization of the seed trade began. Local land races that were produced and maintained by farmers were replaced by seeds that were grown over large geographic distances and that were produced by breeding companies and the seed trading companies. The seed trade started to become regulated and certified, but no formal laws existed (in contrast to other countries).
The modern plant breeding began on the large farms in Eastern Germany at the end of the 18th century. For this reason, many breeding companies were owned by aristocrats who also owned the large farms. Later, however, breeding efforts at public institutions became important, particularly at the Universities of Berlin and Halle because of the infrastructure needed for breeding.
The expeditions of Vavilov
At the same time, Nikolai Vavilov and his colleagues began with their extensive collection trips throughout the world, which led to the establishment of the seed bank in St. Petersburg and to the development of his theory of centers of genetic diversity (Figure 7). Vavilov also founded or took over breeding stations throughout the Soviet Union to utilize the collected material and to create improved crop varieties. There were in total about 200 breeding stations that stored and investigated parts of Vavilov's seed collection.
The first German expedition for plant genetic resources
In 1926 Erwin Baur undertook the first collection trip to Turkey which was explicitly aimed towards collecting new material for the breeding of cereal plants. In 1927, the Kaiser Wilhelm Gesellschaft funded the first institute aimed towards plant breeding in Muencheberg/Mark, close to Berlin. In the same year, Nikolai Vavilov presented his ideas at the V. International Conference of Genetics in Berlin, which was organized by Erwin Baur. His theory of the centers of biological diversity had a great influence on subsequent activities in Germany. The material that Baur collected in Turkey was the founding seed stock of the KWI institute in Muencheberg.
Acquisition of genetic material from the Sovjet Union during the second world war
After the occupation of large parts of Russia and the Ukraine, some leading scientists suggested to the Kaiser-Wilhelm-Gesellschaft (the leading scientific science organisation in Germany, and the predecessor of the Max-Planck-Society) and the government to acquire the seeds of the breeding institutes of the Vavilov organisation in the occupied regions of the Sovjet Union, and to transfer them to the German breeding institutes (Figure 8).

Many Kaiser Wilhelm Institute (KWI) scientists followed the German army behind the lines and collected material from institutions and fields. After the defeat at Stalingrad in early 1943 and the advance of the Red Army, it became clear that the duplicates of the so-called world collection of Vavilov at the Institute in Leningrad would have to be collected from the breeding stations in Ukraine and other occupied territories and brought back to Germany.

One of the most notorious expeditions was organized and conducted by Heinz Bruecher who had studied biology in Tuebingen, became a soldier during the early years of the wars and was then Professor in Jena Thornström and Hossfeld (2002). He was hired by the SS-Ahnenerbe, a scientific branch of the SS to study ancient history and genetics (and other things). He was authorized to lead a 'SS Sammelkommando' (a collection expedition) to collect the ex situ plant material in the Ukraine and Crimea. Because of the growing importance of Lysenkoism (Wikipedia) and the arrest of Vavilov, the genetic resources had not been much of value for the Sovjets and were not moved eastwards beyond the Ural. Therefore the Germans found them relatively untouched.
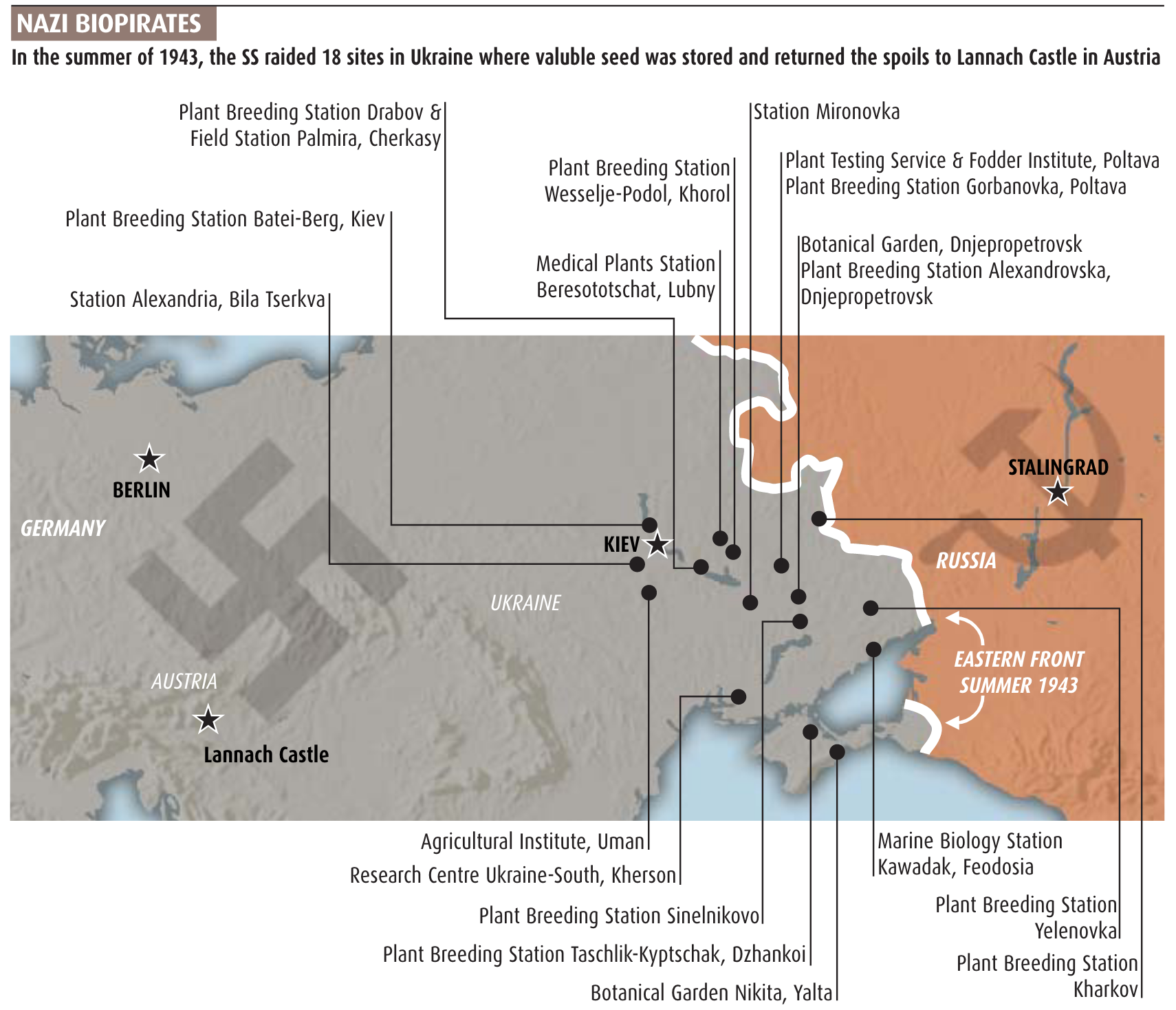
Bruecher was able to collect several thousand samples from the stations and brought them to a SS institute of Plant Genetics at Lannach in Austria, close to the city of Graz. The material was hidden during the end of the war in farms around Lannach and in 1947 brought to West Germany, and its subsequent fate is not clear.
Brücher wrote an extensive report about his experience, which was very informative, in particular with the high quality of scientific research and agricultural practice (and poor leadership of the German occupation) on these agricultural stations (Figure 11).

The genetic resource movement and the establishment of modern genebanks
At the beginning of the 20th century, different people were important for the origin of the genetic resource movement. A good overview was written by R. Pistorius Pistorius (1997).
Germany
In Germany, some plant breeders and geneticists recognized early the importance of the conservation of plant genetic resources.
The two most important plant breeders were Erwin Baur (1875-1933) and Theodor Roemer (1883-1951). Baur formed a 'school' of internationally important crop scientists, that included
- Reinhold von Sengbusch (1898-1987)
- Hans Stubbe (1902-1989)
- Rudolf Schick (1905-1969)
- Hermann Kuckuck (1903-1992)
and who worked in the beginning of their careers at the Kaiser-Wilhelm-Institute (KWI) of Biology in Berlin-Dahlem and at the KWI Institute for Plant Breeding in Muencheberg, close to Berlin. In 1943, during World War II, the KWI of Crop Research was funded close to Vienna. Its first director was Hans Stubbe. At the end of the war the collections from the SS institute in Lannach and the KWI institutes in Vienna and Muencheberg were moved to West and East Germany, where they most likely ended up to a large degree in Braunschweig and in Gatersleben. The move was conducted with the help of plant breeding companies such as Saatzuchtfirma Dippe (Quedlinburg) and the Kleinwanzlebener Saatzucht (today: KWS Group SE).
In 1945, a large proportion of the collection was moved to Gatersleben, close to Quedlinburg. Important collections from the 1930s and 1940s were moved to Gatersleben and the institute was built up rapidly. The institute with its gene bank became then an Institute of the East German Academy of Sciences. After the German reunification, the institute was renamed to the Leibniz-Institute of Plant Genetics and Crop Plant Research (IPK), which hosts now the official German PGR gene bank (Website). The history of the genebank, which is now one of the largest world wide is told by Muntz and Wobus (2012).
Current state of the gene bank at the IPK Gatersleben
It has been estimated that only one third of the taxa present in the gene bank have been subjected to plant breeding efforts Hammer and Gäde (1993). The remaining taxa are land races or wild relatives.
The initial material came from various expeditions conducted in the years between 1935 to 1958:
- Hindukush expedition in 1935
- Travels to Himalaya and Tibet by Von Rauch and Schaefer, 1939
- A collection of Austrian land races from the 1920's and 1930s by E. Mayr
- Expeditions to Ethiopia and Anatolia in 1928 and 1932
- A travel to Spain in 1939
- Expeditions to the Balkans and China in 1941/1942 and 1955/56 by Stubbe.
- Exploration in Italy, 1950
- Expeditions to Iran by Kuckuck in 1952/54
- Expedition to the Mediterranean area in 1958.
Until the German reunification in 1990, further collection expeditions were conducted.
As Table Table 1 shows, there is a substantial number of varieties that were generated by using material from the genebank in Gatersleben.
| Crop | No. of varieties |
|---|---|
| Spring barley | 30 |
| Winter barley | 3 |
| Spring wheat | 1 |
| Winter wheat | 12 |
| Dry soup pea | 2 |
| Fodder pea | 3 |
| Lettuce | 1 |
| Vegetable pea | 4 |
| Total | 56 |
International developments
International developments 1943 to 1959
Henry A. Wallace, the founder of the Pioneer Hi-Bred Company that became powerful with the marketing of hybrid maize, and Raymond Fosdick, the president of the Rockefeller Foundation, decided on an international program that later became famous as the Green Revolution.
In 1943, breeding researchers were sent to Mexico to fund Office of Special Studies, the precursor of the International Maize and Wheat Improvement Centre (CIMMYT). Similar initiatives and institutes were started in other countries, and to a large degree they were funded by the Rockefeller Foundation (http://www.rockefellerfoundation.org).
A rising problem was the internationalization of the seed trade and the agricultural politics. The world was divided into developed and underdeveloped countries. On 16 October 1945, the Food and Agriculture Organisation (FAO) was founded, even before the foundation of the United Nations (UN). In the first years of FAO, seeds were distributed internationally. Later, the collection and preservation of seeds and varieties became more of interest because the danger of a narrow genetic basis was recognized. From 1959 on, collections of plants were initiated. The FAO was given the task
…to do everything possible (…) especially in coordinating plans for plant exploration, in order that collections should be efficiently organized and service, and the proceeds shared equitably among interested parties.
In 1967, the First Technical Conference on Plant Genetic Resources was held. It was based on four premises:
- For advanced cultivars, there exist generally sufficient mechanisms of information and exchange that do not need to be improved much.
- Primitive varieties and wild species are threatened by massive genetic erosion.
- It has to be reacted immediately against this condition, for which there is little time and an international collaboration is required.
- The widely distributed methodological and theoretical knowledge on the collection and conservation has to be collected in a standard treatise for the broad interested public.
The shift changed from the plant introduction and plant exploration to the plant genetic resources. Also the term genetic erosion was coined in this context at the FAO conference in 1967 by Otto Frankel. In this context, Frankel was influenced by Hermann Kuckuck, the German botanist who worked for several years in Iran and Turkey and witnessed the disappearance of the land races of wheat after the introduction of modern high yielding wheat varieties.
This conference marked also the beginning of the discussion on the best type of agriculture and type of conservation: Is it sufficient to conserve most genetic resources ex situ in gene banks and to promote a modern, industrialized agriculture? Or, alternatively, preserve the small-scale agriculture in developing countries where the farmers also manage locally adapted landraces (they were also called primitive varieties) in situ to prevent genetic erosion? The concerns regarding ex situ conservation were expressed by Erna Bennett:
I see no special advantage in conservation in the form of seed apart from the very eminent one of convenience, and I think that attempts to find other merits in the 'steady state' which seed storage represents, seem to come dangerously near to adopting museum concepts. The purpose of conservation is not to capture the present moment of evolutionary time, in which there is no special virtue, but to conserve material so that it will continue to evolve. Such 'continued evolution' could only be possible in in situ collections.
On the other hand, Frankel strongly supported an ex situ conservation system. Given relatively high costs of ex situ conservation, such centers are expected to be located in the developed countries.
To disseminate knowledge about plant genetic resources conservation, the FAO Plant Introduction Newsletter was funded in 1957 and was the most important medium for disseminating information to international breeders. In 1971 it was renamed into Plant Genetic Resources Newsletter5
An important article was written by Jack Harlan 1975 Harlan (1975), in which he outlined the global danger of a loss of genetic variation. To achieve these goals, there have been plans to establish a system of international gene banks under the guidance of FAO or UN because it was realized that the international exchange of seed material using national gene banks would not work.
Pistorius (1997) states that the early efforts of the conservation of PGRs in response to the massive introduction of modern, and in particular hybrid varieties have important characteristics: Conservation and use of PGR were closely linked, because plant breeding companies played an important role. Initially, storage of PGR took place in industrialised countries and was tied to research institutes of plant genetics.
Important consequences were a preference of ex situ over in situ conservation of PGR, and the use of generalist collections. However, it should be noted that discussions of the relative merits of in situ versus ex situ collections continue until today.
The most important results of the 1967 FAO/IPB Conference
Discussions at the conference were quite controversal and they had a political as well as a scientific dimension. The situation in 1967 was characterized by national collection efforts and national genebanks with little communication between them. For this reason, the global conservation of genetic resources did not follow a common plan and was not comprehensive. During the conference it was decided that highest priority is given to the conservation of landraces. Based on the initiative of Sir Otto Frankel who became a leading in the PGR movement in the 1950s, the following principles were agreed upon that can be considered as the dogma of PGR:
Highest priority is given to the collecting and preservation of the widely endangered landraces.
One may differentiate between generalist instead of mission-oriented collecting strategies. A mission-oriented strategy is a collection effort for a particular crop which had to be improved in its particular range of adaptation.
Both strategies have the following characteristics:
- Representativeness through large size of collections.
- Evaluation of the genebank accessions is essential for utilization.
- Preservation by long-term storage.
A goal was to establish a neutral organization unter the umbrella of an international organization that coordinates the conservation of genetic resources. However, the FAO was considered to be too bureaucratic and inefficient. It was decided to establish a system of international gene banks.
The disadvantages of ex situ collections were also discussed:
- Deterioration of collections (loss of germination ability)
- Genetic drift and enrichment of deleterious mutations
- Difficulties in the evaluation of large collections
- Lack of use
- Poor funding
Political problems because of a centralized ex situ storage For these reasons, alternatives to ex situ collections were also considered.
At this time, genecology, i.e. the study of population genetics in the relation to the habitat, was promoted to understand the adaptation of plants to their environment. This field is called molecular ecology today.
The basic assumption of genecology was that adaptation of plants to their environment is polygenic, i.e., it is controlled by multiple co-adapted genes and that selection on multiple genes has occurred throughout historical plant breeding. For this reason it was proposed that selection of individual genes should be reduced in favor of multiple genes, for example in breeding for pathogen resistance. The conservation of plants ex situ would stop the complex evolutionary process involving multiple genes.
This approach was criticized by Frankel, because polygenes are much more difficult to be used in breeding than single genes, and that collection of new genetic resources would require a lot of additional information on local habitats because the polygenes need to be considered in this context. Furthermore, breeders are likely not much interested in locally adapted varieties.
This conflict represents a breeding strategy on farmer's field versus a breeder's plots. Breeders try to isolate useful genes from their genomic neighborhood and introduce them into their own breeding material.
One of the reasons why ultimately ex situ conservation was implemented was that the centralized management of genetic resources is cheaper and more practical.
Another conflict was on the issue of generalized vs. mission-oriented collections. People close to applied plant breeding such as Norman Borlaug advocated the idea of mission-oriented collections because it is assumed that enough overall genetic variation has been collected. In his opinion the bottleneck was the identification and incorporation of useful genetic variation into modern varieties. This argument was countered by the fact that one of the ancestors of the green revolution super rice IR8 (Wikipedia) was a land race from a generalist collection.
The conference concluded with the following results:
- Genetic erosion was recognized and publicized as a problem in breeding
- It was recognized that modern plant breeding requires a steady flow of new germplasm
- A generalist approach was considered to be more successful to prevent genetic erosion
- Genetic variation should be predominately conserved ex situ in gene banks
The ex situ versus in situ conservation debate was not resolved: Are they complementary or alternative strategies?
International developments from 1970 to 1987
Some important dates on the international development: Since 1965, the modern high-input varieties have been introduced world wide. The term 'Green Revolution' has become well known. In 1970, Norman Borlaug was awarded the Peace Nobel Price. In 1971, the World Bank suggested to provide the agricultural research institutes in developing countries with more broad international funds. The Consultative Group on International Agricultural Research (CGIAR) was founded (http://www.cgiar.org). The funding agencies were the private Rockefeller and Ford foundations, the World Bank, and 16 donor countries. The CGIAR headquarter was based at the World Bank in New York. Four institutes that were funded until then by the Rockefeller and Ford Foundations became members of CGIAR: CIMMYT (Mexico), CIAT (Columbia), IITA (Nigeria) and IRRI (Philippines). New institutes were founded: WARDA (Ivory Coast) and CIP (Peru).
![Norman Borlaug Source: Wikipedia. (/img/norman_borlaug.png){#fig-normanborlaug}
The legal construct of CGIAR was difficult to understand. It was an informal network of governments, international organisations and private institutions, co-financed by the world bank, the FAO and the UN developmental program (UNDP). The CGIAR is attached to the UN, but can not be controlled by its member states. It is mainly controlled by the big donor states and organisations in the Northern Countries, some of which have clear commercial interests. One reason for the independence from the UN was that the FAO was considered to be ineffective in achieving the goals.
In the late 1960's and and early 1970's a number of conferences were held that raised the global awareness of plant genetic resources.
In 1969, a FAO panel of experts stated that
- All genetic resources should be made available to all plant breeders immediately and without restriction.
- Genetic variability had to be conserved for future generations in long-term storage with a high level of physical and genetic security.
- In 1972, a panel of American experts published a report on the genetic vulnerability of crops which was highly publicized (NAS report, see below)
In 1973, the UN Environmental Program (UNEP) was decided. In the same year, the International Board for Plant Genetic Resources (IBPGR) was founded that was controlled by the CGIAR and was independent of the UN. IBPGR's mission was to coordinate plant collection and conservation efforts worldwide. It was founded because of a frustration with the FAO to deal with various famines and harvest losses occurring through this period.
A FAO/IPB technical conference drafted an action plan for conserving plant genetic resource IBPGR collected between 1974 and 1984 about 100,000 samples that were stored in the genebanks of the CGIAR institutes. IBPGR was later renamed to IPGRI and is now called Bioversity, with its headquarters in Rome (6).
The action plans suffered from institutional and financial support. The target partners were the national genebanks in the North, and CIMMYT and IRRI genebanks in the developing countries. Little funding came from FAO, because developing countries did not support the initiative. Hunger was highly prevalent in the early 1970s and the developing countries wanted to alleviate immediate problems. Conservation of PGR was considered a long-term activity that should be supported with extra funds.
Public awareness of genetic erosion
In 1970 was the Southern leaf-blight epidemic in the US, and in the same year an outbreak of coffee rust in Brazil, which lead to higher market prices for coffee.
In Asia, modern varieties rapidly replaced land races. The wheat varieties from CIMMYT were grown on an area of 10 Mio. ha in 1964/1965 in India and Pakistan. At end of the 1960s, 95% of the wheat acreage consisted of modern varieties that replaced the landraces. Monocropping of wheat also was increased at the expense of other crops.
A report of the US National Academy of Sciences (NAS) from 1972 came to the following conclusions:
- The market demands uniformity, both by growers and consumers of crops. Uniformity leads to higher vulnerability
- All major crops are affected by this development
- Public and private breeding are affected similarly
- Breeders are in a difficult position: They are criticized from different parties.
For the first time, the report acknowledged the relationship between genetic vulnerability and genetic diversity.
Following solutions were discussed:
- Diversity through time: Rapid turnover of varieties
- Diversity through anticipation: Early warning system for pathogens
- Diversity in reserves: Create numerous advanced breeding lines
- Rapid deployment of genetic diversity: Establish a system for the efficient import of worldwide PGR into the US, which reflect the national interest of the USA.
Establishing an international network of genebanks
In 1972, the advisory board of CGIAR proposed to establish a World Network of Genetical Resources Centers with the following characteristics:
- A coordinating centre: IBPGR (later to become IPGRI and now Bioversity in Rome)
- Establish genebanks in four existing international agricultural research centers: IRRI, CIMMYT, CIAT, and IITA
- Establish genebanks in new international centres: CIP, WARDA, ICRISAT already existed and joined the network. ILCA and ICARDA were later included.
- Establish new 'regional' centres in Vavilovian centres of crop diversity
In the 1980s, the IBPGR established networks with national governments (donors) to catalyze research and initiate collection programs. IBPGR produced achievements in advice and support, but did not distribute funds.
From the Mid 1970's to the early 1980's, the regional genebanks for local crops were set up, for example the quinoa gene bank in the Andean highlands. However, the regional gene banks were not very successful (particular in the utilization of the material) due to political problems. The government and administrations often had no previous experience with gene banks, and the countries were not willing to exchange material.
National organizations such as the German Gesellschaft fuer Technische Zusammenarbeit (GTZ)7 helped countries to establish gene banks, such as the Ethiopian national genebank in Addis Abeba.
7 GTZ has been renamed to GIZ, http://www.giz.de
How plant genetic resources became the center of a global political conflict
In the 1980s political problems arose between the Northern and Southern countries because only 15% of the samples that were nearly exclusively collected in the developing countries ended up in the national gene banks of the developing countries. Most other accessions were stored in the national gene banks of Northern countries or in the CGIAR gene banks that had an uncertain legal status (especially with respect to ownership).
There were also political gene embargoes. In the 1980s, the US did not send seeds to Libya, the Soviet Union and to Nicaragua.
Two NGO organisations that were run by very successful activists turned the conservation and exchange of PGR into a global political issue. These organizations, RAFI and GRAIN stressed in particular the implication of the current situation for poor and developing countries.
The key arguments were as follows:
- PGR are useful for companies
- In particular with genetic engineering: By patenting a strong protection of crops can be achieved.
- Genetic resources come under the control of companies
- CGIAR is strongly influenced by donors with commercial interests.
The different arguments are summarized in the monography of Pistorius (Figure 13).
The Convention of Biological Diversity (CBD)
In the wake of this controversy, a discussion arose as to whether PGR are freely exchanged. The questions of ownership and the legal status of PGR had to be answered. This was achieved with the Convention of Biological Diversity (CBD) and the subsequent International Treaty of Plant genetic resources. The CBD was signed at the UN Environmental Conference in Rio de Janeiro 1992 and implemented in December 1993. The signing states get the international rights over their biological resources, with the following limitations: Only genetic resources for which they are country of origin can be controlled or states that have acquired genetic resources after 1993 (or the year they signed the CBD). Since most CGIAR ex situ collections were established before 1993, they are (legally) not covered by the CBD!
The legalization of the natural environments was advanced by two new developments:
- The environmental program of the United Nations (UNEP)
- The GATT treaty
From these efforts, the Convention of Biological Diversity (CBD) originated. The key aspect is that the countries are responsible themselves for managing and utilizing their biological diversity. Only material that was collected after 1993 is part of the CBD and needs to be sent out with the Standard material transfer agreement. However, some countries try to push the date back to before 1993. The exchange of genetic material is now in the responsibility of each country holding the resources.
Key concepts
| \(\square\) Convention on biological diversity |
| \(\square\) Consultative Group of International Agricultural Research (CGIAR) |
| \(\square\) Generalist vs. mission-oriented collection strategy |
Summary
- The movement of to preserve plant genetic resources is closely tied to the desire to protect genetic resources.
- In Germany, very early a system of public institutes to collect and utilize plant genetic resources were created.
- The Genebank in Gatersleben has become one of the largest gene banks in the world, thanks to the effort of many collection expeditions.
- After the second world war, the conservation of genetic resources has become an international effort.
- Because of the internationalization of seed companies and the emerging North-South conflict, the arena has become highly politiced.
- The Convention on Biological Diversity and the GATT treatment regulate the collection, exchange of genetic resources and any intellectual property derived from them.
Further reading
A good and concise summary of historical and political aspects of the genetic resources movement is by Robin Pistorius, Scientists, plants and politics - A History of the Plant Genetic Resources Movement. (1997) PDF available at https://www.bioversityinternational.org/e-library/publications/detail/scientists-plants-and-politics/
Review and discussion questions
- Why is the study of the historical distribution of crops relevant for political and scientific aspects of plant genetic resource characterization and utilization?
- What were the key international developments and events in the history of the plant genetic resource movement?
- Which organisations and institutions originated from these developments, and what is their task?
- What are the key conflict lines that arose during the development of the current system of plant genetic resource conservation?
- What was the impact of the Green Revolution on plant genetic diversity?
Problems
- Go to GBIF and download data.
- ETc